Вернуться на страницу ежегодника Следующая статья
II. МОЗАИКА СОЦИАЛЬНОЙ ЭВОЛЮЦИИ
Эволюция человека: между конкуренцией и сотрудничеством* (Скачать pdf)
Марина Львовна Бутовская, Российский государственный гуманитарный университет; Институт этнологии и антропологии РАН
В статье обсуждаются вопросы, связанные с эволюцией социального поведения человека, и рассмотрены факты в подтверждение теории генно-культурной коэволюции. Показаны сходство и различие моделей конкуренции и кооперации у человека и нечеловеческих приматов. На примере детей и подростков из четырех традиционных африканских обществ проанализирован дележ ограниченными ресурсами и показана роль индивидуальных факторов, социального воспитания и культурных норм при решении дилемм дележа в отношении друзей и незнакомцев. Показано, что малые и большие общества имеют значительно больше сходства по моделям кооперации, нежели считалось ранее.
Ключевые слова: эволюция социальности, групповой отбор, кооперация, дележ ограниченными ресурсами, малые и большие общества.
Человеческое общество как продукт эволюции
Место человека в эволюционной картине мира – одна из наиболее обсуждаемых тем в антропологии. В наше время тезис о двойственной, биосоциальной природе человека разделяется практически всеми специалистами (Бутовская, Файнберг 1993; Бутовская, Ростовцева 2021; Вааль 2014; 2020; 2021; Hrdy 2009; 2016; Tomasello 2020а; 2020b; Burkart et al. 2018; Gintis et al. 2019; Sánchez‐Villagra, van Schaik 2019; Pandit et al. 2020). Причина тому ‒ фантастические достижения в области антропологии, этнографии, археологии, этологии человека, приматологии, палеогенетики, нейропсихологии и целого ряда других интенсивно развивающихся дисциплин, изучающих природу человека и позволяющих не только анализировать особенности функционирования социальных объединений у разных видов приматов и в разных современных человеческих обществах, но и судить о характере социальных связей в обществах людей далекого прошлого (Бутовская 1987; Sánchez‐Villagra, van Schaik 2019; Pandit et al. 2020; Frayer 2019; Sikora et al. 2017; Santos et al. 2021; Zwir et al. 2021).
Социальное поведение человека ‒ такой же продукт эволюции, как и его морфология и физиология. В этом контексте проявления конкуренции и кооперации следует рассматривать в свете комплекса факторов, включая социальные, когнитивные, личностные, онтогенетические и биологические. Для понимания человеческой социальности и социального поведения важно учитывать теорию генно-культурной коэволюции. В рамках этой теории генетические характеристики формируются под влиянием средовых факторов (экология) и в равной мере под действием факторов социальных. Между врожденными свойствами и приобретаемыми в процессе онтогенеза (социализации) характеристиками поведения, таким образом, существует обратная связь. Поскольку личностные характеристики отбираются под давлением социума, и в то же время характеристики социума определяются в существенной мере составом этого социума, культурная эволюция может оказывать непосредственное воздействие на социальную и физическую среду, изменяя их, а это приводит к изменениям генофонда. Показательным примером выступает переход к приготовлению пищи на огне. Распространение этой традиции стало мощным селективным фактором и повлекло за собой перестройку кишечного тракта, желудка, зубной системы у наших палеолитических предков (Wrangham 2017). А сокращение энергетических затрат на переваривание пищи освободило дополнительную энергию для развития и функционирования мозга, что в свою очередь привело к прогрессивному развитию интеллектуальных способностей.
Как у любого другого общественного вида животных, конкуренция и кооперация выступают в неразрывной связи в качестве важнейших проявлений человеческой социальности. Эта связь существует на всех уровнях социальной сложности, начиная с диадных взаимодействий в пределах группы и заканчивая межгрупповыми и надгрупповыми уровнями организации. В течение десятилетий в эволюционной биологии превалировали социобиологические подходы, в рамках которых эволюция альтруизма мыслилась исключительно в русле понятий индивидуального отбора (Bradley 1999; Sillen-Tullberg, Bryant 1983; Wilson 1978), однако к настоящему моменту все больше специалистов (прежде всего антропологов и этологов) обращаются к понятиям группового отбора и, прежде всего, приводят доказательства его исключительной роли в эволюции человека и общества (Бутовская, Ростовцева 2021; Бутовская и др. 2021; Henrich 2016; Newson, Richerson 2018; Richerson, Boyd 2005; Nowak 2006; Apicella, Silk 2019; Smith et al. 2018; Waring, Wood 2021). Ни один другой вид на нашей планете не зависит столь сильно от социальной (культурной) информации, как человек. Наш вид ультрасоциален. Мозг человека, его геном и биология в течение сотен тысяч лет формировались в контексте тесной взаимосвязи культурной и биологической эволюции, и именно культурный отбор открыл новые горизонты для уникальных генно-культурных трансформаций (Henrich, Muthukrishna 2021).
Сотрудничество в человеческом обществе происходит постоянно и повсеместно, оно столь привычно, что мы перестали замечать его. Современные исследования в области приматологии примиряют сторонников и противников идеи об уникальности феномена сотрудничества у человека. Полевые и лабораторные данные показывают, что внутривидовая кооперация широко распространена в животном мире и не ограничивается только человекообразными обезьянами. Вместе с тем только у человека она достигает беспрецедентных масштабов, и сети кооперации могут включать не только родственников, друзей и знакомых, но и незнакомцев, в том числе и представителей другой культуры.
Сотрудничество и групповой отбор
Среди исследователей эволюции социальности не прекращаются споры о том, как и почему особи одного вида помогают друг другу, и делается акцент на уникальную роль культуры как фактора эволюции у человека (Laland et al. 1995; Nowak 2006; Richerson et al. 2016; Бутовская, Ростовцева 2021). Помощь является дорогостоящим продуктом и часто не только требует энергетических затрат, но и сопряжена с рисками для здоровья и даже самой жизни. Впрочем, из этого не следует, что теория генно-культурной коэволюции не работает применительно к животным сообществам. Благодаря культурно-обусловленным миграциям и репродуктивной ассортативности культура формирует специфические черты популяционной генетической структуры у животных и определяет внутрипопуляционное разнообразие у социальных видов (Whitehead et al. 2019).
Существенный шаг в понимании роли группового отбора был сделан М. Новаком, предложившим объяснение механизмов эволюции кооперации на индивидуальном и групповом уровнях (Nowak 2006). Дальнейшее развитие этих взглядов было представлено в публикации К. Апицеллы и Дж. Силк (Apicella, Silk 2019). Авторы этой работы показали, что в череде поколений обманщики, получая индивидуальные преимущества, все же постепенно вымываются из популяции на всех уровнях: родственного отбора, прямой реципрокности, группового отбора и также при выборе полового партнера. При этом ведущим фактором выступает ассортативность. Как бы ловко ни маскировались обманщики, их все же вычисляют окружающие, после чего стараются от них дистанцироваться и передавать соответствующую предупреждающую информацию окружающим. В результате соответствующих социальных перестановок и реструктуризации социальных связей кооператоры (люди, оказывающие помощь другим) постепенно объединяются друг с другом, и такие объединения оказываются более сплоченными перед лицом различных угроз и внешних вызовов, а их члены лучше выживают в условиях межгрупповой конкуренции.
Выдающиеся теоретики прошлого Томас Гоббс и Жан-Жак Руссо, невзирая на принципиально различные взгляды на природу человека и общество далекого прошлого (злость и алчность, война всех против всех в первом случае, и представление о благородном дикаре и природном альтруизме во втором), полагали, что на заре человеческой истории люди не мыслили себя как часть социума и не сообразовывали свои действия в соответствии с определенными социальными ролями, ими двигали лишь индивидуальные мотивы. Однако сегодня мы знаем, что ближайшие родственники человека, современные человекообразные обезьяны (шимпанзе, бонобо, гориллы) ‒ в высшей степени социальные животные (Вааль 2020; 2021; Бутовская, Ростовцева 2021). У этих видов имеется отчетливая социальная структура, более иерархичная, с полигамными чертами у горилл, с иерархией взрослых самцов-резидентов у шимпанзе и эгалитарно организованная с доминированием коалиций взрослых самок и статусными привилегиями их половозрелых сыновей у бонобо (Lewis et al. 2021; Ishizuka et al. 2020; Samuni et al. 2021). У шимпанзе и бонобо, имеющих сходную социальную структуру обществ с группами, распадающимися и объединяющимися во времени (fission-fusion grouping) (Butovskaya 2020), особи не просто распознают знакомых и незнакомых представителей своего вида, но предпочитают визуально следить за знакомыми представителями доминирующего пола (за самцами – у шимпанзе, за самками – у бонобо) (Lewis et al. 2021). Очевидно, что различия в социоэкологии этих видов оказали влияние на способности к непосредственному сотрудничеству (кооперацию), различия в направленности социального внимания и системе оповещения сородичей об опасности.
Многие авторы говорят о большей просоциальности бонобо по сравнению с шимпанзе (Вааль 2020; 2021; Nolte, Call 2021). В недавних экспериментах с шимпанзе и бонобо проверяли готовность одного из членов пары оказывать помощь, передавая другому орудия, необходимые для достижения цели, в двух вариантах сценариев: а) помощник не получал от таких действий никакой выгоды; б) помощник получал награду при условии правильного выбора орудия и передачи его партнеру (Nolte, Call 2021). В парах шимпанзе помощь оказывалась в единичных случаях и только от матери к дочери, тогда как у бонобо периодически помощь оказывалась в обоих сценариях, но только в парах самок, в парах самец ‒ самка помощь отсутствовала. Эти эксперименты свидетельствуют о том, что бонобо (самки) лучше понимают социальные проблемы и более склонны к сотрудничеству (Ibid.), ясно также, что просоциальность выражена у самок этого вида значительно лучше, чем у самцов.
Оба вида (шимпанзе и бонобо) способны к передаче важной социальной информации членам группы, но у шимпанзе система оповещения об опасности более эффективна (Girard-Buttoz et al. 2020), что логично связать с большей напряженностью в отношениях между соседними сообществами у шимпанзе и агрессивными рейдами самцов с летальными последствиями для животных их соседней группы (Wilson et al. 2014). У шимпанзе вероятность участия в межгрупповых взаимодействиях (преимущественно агонистических) возрастает с числом участников такой коалиции, с ростом числа родственников по материнской линии и друзей, входящих в коалицию (Samuni et al. 2021). Эти данные указывают на способность шимпанзе понимать сущностную сторону социальных отношений и оценивать качественный эффект социальных связей на групповом уровне. Самуни с соавторами подчеркивают очевидную значимость социальных связей у шимпанзе и их интегральную роль для развития сотрудничества. Важнейший вывод из этой работы ‒ доказательство взаимосвязи между прочными и устойчивыми социальными отношениями партнеров внутри группы и совершением рискованных и опасных коллективных действий с выгодой для группы. Подчеркнем, что ранее подобное поведение считалось исключительно прерогативой человека, но с учетом накопившихся к настоящему времени фактов к этому тезису нужно относиться со всей осторожностью.
Нет ни малейшего сомнения в том, что предки человека, равно как и все виды гоминин, были социальными (Бутовская, Файнберг 1993; Вааль 2020; 2021). Их общества обладали выраженной социальной структурой и иерархией, а статус особей определялся с учетом их личных качеств, родственных связей, возраста и пола. По-видимому, общие предки человека и шимпанзе обладали как минимум базовыми способностями к сотрудничеству, но это не означает, что конкуренция отсутствовала как на внутри-, так и на межгрупповом уровне. Напротив, именно конкуренция между группами может рассматриваться в качестве ведущего стимула к развитию сотрудничества на уровне группы (Samuni et al. 2021) и кооперативной мотивации между ее членами (Girard-Buttoz et al. 2020).
От обществ нечеловеческих приматов к человеческим обществам (простые и сложные общества)
Групповой образ жизни – важнейшая адаптация социальных видов, дающая особям очевидные преимущества по сравнению с одиночным существованием (Alexander 1974; Бутовская, Файнберг 1993; Бутовская, Ростовцева 2021). Однако у членов одного общества регулярно возникают конфликты интересов в вопросах различных ограниченных ресурсов, чреватые антагонизмом на уровне конкретных пар, иерархия доминирования стабилизирует отношения в пределах общества и снижает вероятность вспышек физической агрессии в этих условиях. Статус особей в обществе человекообразных обезьян чаще всего зависит от способности выиграть драку и умения объединяться в коалиции с другими особями для достижения более высокого статуса или защиты уже имеющегося. В отличие от обществ животных, статус в человеческих обществах чаще оценивают с позиций вовлеченности человека во власть (в какой мере он может оказывать влияние на других, не прибегая к открытому насилию), с учетом его родственных и дружественных связей, организаторских способностей, умения контролировать окружающих и манипулировать ими. Признание центральной роли культуры в человеческом обществе позволяет рассматривать социальный статус человека с эволюционных позиций не просто как продолжение иерархий доминирования у приматов, но и как феномен, завязанный на развитие когнитивных способностей (Henrich, Gil-White 2001). Воплощением статуса может быть простое доминирование или такое более сложное явление, как престиж. Оба варианта описаны в человеческих обществах, но их роли существенно отличаются, причем второй вариант значительно облегчает культурную трансформацию и передачу новых идей в обществе.
Чтобы пролить свет на загадку человеческой ультрасоциальности, нуж-но не только обратить внимание на широту и интенсивность человеческого сотрудничества, но и задаться вопросом о том, почему проявления сотрудничества существенно варьируются в разных обществах – как в географической, так и в исторической перспективе.
Дж. Хенрич и М. Музукришна (Henrich, Muthukrishna 2021) утверждают, что эволюционный подход, ориентированный на признание роли генно-культурной коэволюции в формировании когнитивных различий и кросскультурных различий в психологии, заслуживает особого внимания. В ряд таких эволюционных механизмов, позволяющих объяснить феномен человеческого сотрудничества, авторы ставят родство, реципрокность в выборе партнера, репутацию, нормативы и коллективные средства подавления-наказания. Естественный отбор формирует психологические установки на общение с другими людьми и мотивацию к заботе и сотрудничеству с ними (Henrich, Muthukrishna 2021). Культурные нормы сотрудничества усваиваются детьми и подростками в процессе интеграции в социальные сети конкретного общества. Важно отметить, что неолитическая революция и потребности производящей экономики не только привели к усилению межгрупповой конкуренции, но и усилили рост внутригруппового единства и сотрудничества.
Предрасположенность к альтруизму может фиксироваться на генетическом уровне, и группы с большим количеством альтруистов оказываются более сплоченными, кооперативными и вместе с тем более конкурентными в направлении других групп (Czárán, Aanen 2016; Gintis et al. 2019). Способность проявлять предпочтения в отношении других имеет решающее значение для развития кооперации в больших обществах, там, где большие группы генетически неродственных людей находятся в повседневном взаимодействии друг с другом. Развитие базовых механизмов в основе избирательного предпочтения других обеспечило основу для развития эгалитаризма и парохиализма у человека (Fehr et al. 2008).
Дележ ограниченными ресурсами у детей и подростков в традиционных африканских обществах
Хотя дележ в ограниченном виде присутствует у других приматов, он чаще всего ограничен отношениями «мать – детеныш» и носит однонаправленный характер или же представляет собой пассивный дележ, когда реципиент берет пищу рядом с другой особью, подбирает оброненные высокоранговой особью кусочки лакомства (например, мяса). Только у человека дележ носит целенаправленный и двусторонний характер. Люди обмениваются пищей, делятся с другими (мать – ребенок, сверстники, партнеры). Только у человека дележ пищи практикуется на внутри- и меж-групповом уровне и включает неродственных индивидов, охватывает как знакомых, так и незнакомых индивидов.
Корни дележа восходят к сообществам ранних охотников-собирателей. Наши предки столкнулись с беспрецедентной ситуацией, выбрав для жизни нишу охотника и собирателя и конкурируя как с хищниками за животную пищу, так и с другими видами приматов за растительные ресурсы. Ниша охотника-собирателя связана с риском и неопределенностью добычи пищи. Этот фактор являлся одним из решающих при формировании адаптации к широкомасштабному дележу пищи у человека.
Современные исследователи часто используют экономические игры и их элементы как инструмент для исследования механизмов кооперации и альтруистического поведения у человека. Э. Фер и его коллеги (Fehr et al. 2008) одними из первых разработали серию экспериментов с дележом ограниченными ресурсами для проверки индивидуальных различий в тактиках дележа (установок на просоциальность, зависть и истинный альтруизм). Анализируя результаты экспериментов с детьми в возрасте от трех до восьми лет, эти авторы показали, что с возрастом повышается эгалитаризм в просоциальной игре и игре на справедливость независимо от на-правленности дележа «свой – чужой». В случае же с альтруистическим дележом дети демонстрируют парохиальные установки. В экспериментах Фера принимали участие дети из европейских постиндустриальных обществ.
В какой мере культурная специфика и экономика влияют на выбор стратегий дележа в направлении друзей и незнакомых сверстников, мы проверили на выборке детей и подростков из четырех традиционных обществ Восточной Африки (хадза, ираку, меру, хая). Общая выборка составила 1075 человек, средний возраст испытуемых – 13,0 ± 3,0 г. Материалы были собраны нами с 2013 по 2021 г. Более подробно описание выборок и методов см.: Бутовская и др. 2021; Butovskaya, Burkova, Karelin 2020; Butovskaya, Karelin et al. 2020).
Как уже говорилось выше, нами применялся экспериментальный метод оценки просоциальных
ориентаций детей и подростков, исходно разработанных Э. Фером (Fehr et al. 2008). В играх участник принимал решения относительно дележа
конфет с воображаемым партнером. Все участники играли в три игры: просоциальную,
игру на зависть и истинный
дележ, и каждая игра была в два кона: 1) в направлении друга; 2) в направлении незнакомого сверстника.
Распределение респондентов, предпочитающих альтруистские, просоциальные, смешанные и эгоистические стратегии дележа ограниченными ресурсами для всей выборки представлено на рис. Бросается в глаза тот факт, что в целом подавляющий процент детей и подростков ведут себя просоциально и альтруистично в отношении реципиентов вне зависимости от того, знакомые ли это сверстники (друзья – 78,51 %; незнакомцы – 72,59 %). Хотя все же больше детей демонстрируют истинный альтруизм в отношении друзей (рис. А, Б). Соответственно, несколько меньше детей проявили себя жадными в отношении друзей по сравнению с незнакомцами. Частная корреляция между установками в отношении друзей и незнакомцев с контролем по полу, возрасту и этническому происхождению оказалась положительной и высокодостоверной (r = 0,45, p = 0,0001). Хотя определенная парохиальность при распределении ограниченных ресурсов имела место, все же нужно отметить, что мотивация дележа проявлялась у большинства детей, и степень знакомства между актором и реципиентом не играла решающей роли. Дети с выраженной альтруистичной мотивацией вели себя практически идентично в ситуациях дележа с друзьями и незнакомцами, а жадины не хотели делиться вовсе.
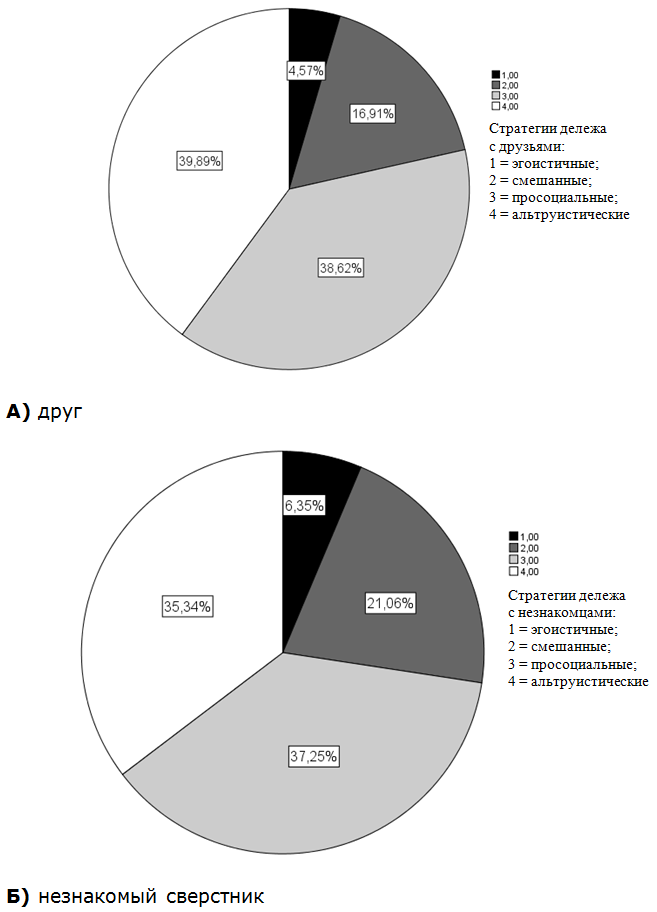
Рис. Распределение детей из общей выборки, практикующих альтруистские, просоциальные, смешанные и эгоистические стра-тегии дележа в направлении: А) друга; Б) незнакомого сверстника
На следующем этапе мы рассмотрели распределение четырех типов стратегий в направлении друзей и незнакомцев в четырех африканских сообществах по отдельности (табл.). Наибольший процент детей, ориентированных на эгоистические стратегии при дележе ограниченными ресурсами по отношению к друзьям, отмечен у хадза, а наименьший – у меру. Меньше всего детей, практикующих смешанные стратегии в отношении друзей, отмечено у иракв, а больше всего у хая. Наибольший процент детей, практикующих просоциальные стратегии в направлении друзей, выявлен у меру, а наименьший – у хая. Наконец, наибольший процент альтруистов в отношении дележа с друзьями отмечен у иракв и хая, а наименьший – у меру.
Табл. Различия в распределении респондентов, практикующих эгоистические, смешанные, просоциальные и альтруистические стратегии в направлении друзей и незнакомых сверстников в четырех африканских обществах
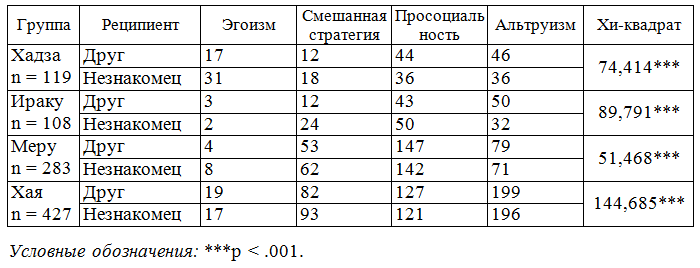
Наше исследование показало, что дружба играет важную интегрирующую роль в развитии просоциальности и альтруизма у детей и подростков в традиционных африканских обществах. Недавние исследования других авторов, проведенные на европейской выборке, дают сходные выводы (Gȕroglu et al. 2014). Выявлены культурные различия в плане проявления эгоизма, просоциальности и альтруизма в направлении друзей и незнакомцев: 1) минимально выраженный альтруизм в отношении друзей – у хадза; 2) максимально выраженный альтруизм в отношении незнакомцев – у хая. В качестве объяснения можно предположить, что хадза – небольшая по размеру группа, и все ее члены знакомы друг с другом. Присваивающая экономика предполагает дележ пищи в случае большого ее количества и толерантна к утаиванию небольших порций лакомой пищи (меда); хая – одна из крупнейших этнических групп. Отмечается компактное ее проживание в сельской местности. Потенциально любой незнакомец в этом случае оказывается также хая. Дележ с ним, таким образом, адаптивен и способствует интеграции группы. Пол играл менее значимую роль в практике дележа ограниченными ресурсами, чем этнос. Однако в целом девочки вели себя более просоциально и альтруистично по отношению к незнакомцам, чем мальчики. Возраст выступает существенно важным фактором, определяющим стратегии дележа во всех изученных этносах в отношении как друзей, так и незнакомцев. В отношении друзей пик эгоистических решений приходится на разный возраст у разных этносов (9–10 лет у хадза и 15 лет у хая), а динамика принятия альтруистических решений показала выраженные пики в нескольких возрастах (11 и 16 лет у меру). В отношении незнакомцев пик эгоистических решений приходился на 17 лет у хая, а альтруистических решений – на 14 лет у меру, 16 лет у хая, 17 лет у хадза. Предпочтительное распределение ресурсов в пользу друзей (по сравнению с незнакомцами) в подростковом возрасте может быть универсальной чертой человеческого социального поведения, проявляющейся в малых и больших обществах. Люди, практикующие эгоистические стратегии распределения ресурсов, оценивали себя как менее счастливых по сравнению с более альтруистическими сверстниками. У хадза стратегии дележа ограниченными ресурсами не зависели от возраста. У ираку с возрастом дети становились менее альтруистичными, но более эгалитарными по отношению к друзьям. В отношении незнакомцев их решения с возрастом стали более эгоистичными. Наши данные свидетельствуют о том, что, в отличие от хадза (охотники-собиратели), у земледельцев и агропасторалистов проявления парохиализма с возрастом становились более выраженными. Эти данные указывают на важную специфику обществ с производящей экономикой – формирование сплоченных групп, ориентированных на внутригрупповую кооперацию в противопоставление чужакам.
Крупномасштабная кооперация в малых обществах возможна
Как уже говорилось выше, в современной антропологии термины «дикость» и «варварство» рассматриваются как оскорбительные, и в научной литературе их называют малыми (или простыми) обществами. Малые общества не превышают по размеру нескольких тысяч человек, занимаются охотой и собирательством, а также скотоводством или простым мотыжным земледелием. Их поселения сельского типа. Малые общества могут проживать на большой территории, как, например, народы Крайнего Севера – чукчи, эскимосы, ненцы – или кочевые и полукочевые народы Африки – бедуины, масаи, датога, ираку. Такие общества не строят городов, не имеют централизованных правительств и сложных экономических отношений. Важным моментом, характеризующим малые общества, является специфика ролевых отношений. В таких обществах практически все знакомы со всеми, и в течение жизни взаимодействуют с теми же индивидами во всех возможных социальных контекстах. В условиях малых обществ каждый человек связан сложной системой социальных обязательств с другими, все его действия повышают или, напротив, снижают престиж его семьи, линиджа и клана, а порой также друзей и соседей.
Фундаментальная загадка человеческой эволюции связана с вопросом о развитии механизмов сотрудничества с генетически не связанными друг с другом незнакомцами. Одним из предлагаемых объяснений является отбор на уровне группы культурно дифференцированных популяций. В недавней работе К. Хэндли и С. Мэтью (Handley, Mathew 2020) проверяется важнейшее предсказание теории отбора культурных групп и предлагается оценка готовности к сотрудничеству между людьми из разных групп с уче-том степени культурного сходства между этими группами. Авторы задокументировали нормативные убеждения и предрасположенность к сотрудничеству 759 человек из девяти кланов, входящих в четыре скотоводческие этнические группы Кении – туркана, самбуру, рендилле и борана. И обнаружили, что сотрудничество между группами зависит от их культурного сходства. Следовательно, делают выводы эти авторы, нормы сотрудничества в данных обществах эволюционировали под влиянием груп-пового отбора на культурные различия. Такой отбор, влияющий на эволюционную историю человека, позволяет объяснить, почему мы охотно сотрудничаем с неродственными и незнакомыми людьми и почему беспрецедентная кооперативная гибкость людей тем не менее демонстрирует выраженную парохиальность (то есть ограничена рамками культурной группы).
В литературе широко распространено мнение о том, что в условиях малых обществ слабо развита централизация, а кооперация носит ограниченный локальный характер (Boehm 2012). Но такая точка зрения не соответствует действительности (Glowacki, Lew-Levy 2022). Хотя кооперация в условиях малых обществ и осуществляется преимущественно между родственниками и друзьями, крупномасштабная кооперация в этих условиях возможна и реально практикуется, причем она может носить экстенсивный характер (Там же). Данные кросс-культурных исследований по ныне живущим охотникам-собирателям показывают, что эти общества способны осуществлять не только широкомасштабную кооперацию, но и организованные действия, участие в которых принимают десятки и даже сотни людей, и нет причин полагать, что в палеолитическом прошлом дела могли обстоять иначе (Singh, Glowacki 2021). Сотни неродственных индивидов могут кооперироваться, чтобы обеспечить режим огненного пала при необходимости, создание многокилометровых заграждений для загонной охоты или ведение военных действий в направлении вражеского сообщества (Bird et al. 2020; Mathew, Boyd 2011; Glowacki, Wrangham 2015).
Общества постоянно балансируют между конфликтом и кооперацией
Как малые, так и большие общества постоянно предпринимают шаги для поддержания внутреннего равновесия, стараясь находить пути для преодоления постоянно возникающих внутренних конфликтов и акцентируясь на внутригрупповой кооперации. Однако в малых обществах эти тенденции проявляются более отчетливо. Универсальным можно считать конфликт между младшим и старшим поколениями, по-разному приходящими к известности и власти (напористость и успешность в военных кампаниях в первом случае и экономическая успешность, обеспеченность, организаторские способности, социальные связи, установленные на основе дружеских и партнерских договоренностей – во втором) (Weissner 2019: 225). Потенциальный конфликт кроется в структуре систем с выборным лидерством, поскольку многие амбициозные молодые мужчины используют возникающие внутригрупповые конфликты и размолвки для формирования собственных политических единиц и выхода из исходной группы. Конфликт в условиях малых обществ, регулярно практикующих рейды в направлении соседних групп, также возникает между верностью братьям по оружию (сражавшимся совместно против группы) и лояльностью к родственникам брачных партеров из враждебных групп (Choi, Bowles 2007). Потенциальный конфликт может также возникать между приверженностью воинскому братству, с одной стороны, и собственным женам и другим родственникам, негативно настроенным в отношении военных действий, – с другой (Rodseth 2012). Все вышеперечисленные фракционные конфликты и противоречия могут привести к смене культурных правил, норм и моральных стандартов войны и мира (Weissner 2019).
В силу того факта, что в условиях малых обществ редко накапливаются излишки пищи, первостепенное значение приобретают неформальные институты, распределяющие риски между отдельными индивидами, семьями и сообществами (Glowacki, Lew-Levy 2022). Для большинства охотников-собирателей центральную роль играет распределение добычи, принесенной охотником, между всеми обитателями лагеря (Butovskaya, Marczak et al. 2020). Важной социальной адаптацией, нейтрализующей риски продолжительного голода, выступают сети обмена по родственным и другим обязательствам, такие как hxaro у бушменов (Wiessner 2002), система дарения, osotua, у масаев (Aktipis et al. 2016), дружеские связи у датога, масаев и других скотоводов (Cronk, Aktipis 2021; личные наблюдения автора), право на доступ к воде, охотничьим и пастушеским угодьям, основанные на родстве и свойстве клановой принадлежности или членстве в соседской общине (Glowaski 2020; Glowacki, Lew-Levy 2022).
Кооперация в человеческом обществе зависит
от целого ряда факторов. Это и культурные традиции в целом, непосредственные условия, в которых оказываются люди, наличие внешнего прессинга со стороны враждебно настроенных
соседей, экономических показателей домохозяйства тех, от кого ожидают акты жертвования
ресурсами. Однако в пределах одного и того же общества четко прослеживается индивидуальная
изменчивость по этому признаку. Такая вариабельность связана с врожденными характеристиками
индивида (в том числе генетической предрасположенностью) и особенностями социализации.
Важным фактором, влияющим на кооперативное поведение, выступает пол. Одно из последних
наших исследований показывает, что важными факторами, влияющими на кооперативные
установки человека, являются порядок рождения, воспитание в многодетной семье (Ростовцева
и др. 2020). Причем для мужчин более значимым оказывается фактор многодетности,
а для женщин – порядок рождения. Как было показано в нашем недавнем экспериментальном
исследовании на бурятских студентах, молодые мужчины из многодетных семей (≥ 4 ребенка
в семье) были более склонны проявлять кооперацию в групповых взаимодействиях. Юноши,
выросшие в семье, где являлись единственным ребенком, оказались бóльшими эгоистами.
Этот факт заслуживает внимания в силу общемировой тенденции к однодетным семьям.
Женщины, являвшиеся первым ребенком в семье (старшая сестра), были наименее кооперативны
в парных взаимодействиях с другими девушками. Последние наблюдения говорят о том,
что старшие девочки в бурятских семьях привыкают командовать и склонны реже идти
на компромиссы, чем младшие по счету дети. В какой мере этот эффект можно экстраполировать
на другие культуры – еще предстоит выяснить
в будущем.
Библиография
Бутовская М. Л. 1987. Эволюция группового поведения приматов как предпосылки антропогенеза. Советская этнография 1: 52–69.
Бутовская М. Л., Дронова Д. А., Карелин Д. В. 2021. Дележ ограниченными ресурсами у детей и подростков охотников-собирателей (хадза) и земледельцев (ираку) в контексте представлений об эволюционных основах альтруизма у человека. Stratum Plus Journal 1: 17–37.
Бутовская М. Л., Ростовцева В. В. 2021. Эволюция альтруизма и кооперации человека: Биосоциальная перспектива. Москва: URSS.
Бутовская М. Л., Файнберг Л. А. 1993. У истоков человеческого общества. М.: Наука.
Вааль Ф. Д. 2014. Истоки морали. В поисках человеческого у приматов. М.: Альпина Паблишер.
Вааль Ф. Д. 2020. Последнее объятие Мамы: Чему нас учат эмоции животных. М.: Альпина Паблишер.
Вааль Ф. Д. 2021. Наша внутренняя обезьяна: Двойственная природа человека. М.: Альпина Паблишер.
Ростовцева В. В., Бутовская М. Л., Мезенцева А. А., Дашиева Н. Б. 2020. Влияние числа сиблингов и очередности рождения на индивидуальную кооперативность во взрослом возрасте: экспериментальное исследование среди бурят. Этнографическое обозрение 5: 162–184.
Aktipis A., De Aguiar R., Flaherty A., Iyer P., Sonkoi D., Cronk L. 2016. Cooperation in an Uncertain World: for the Maasai of East Africa, Need-based Transfers Outperform Account-keeping in Volatile Environments. Human Ecology 44: 353–364.
Alexander R. D. 1974. The Evolution of Social Behavior. Annual Review of Ecology and Systematics 5(1): 325–383.
Apicella C. L., Silk J. B. 2019. The Evolution of Human Cooperation. Current Biology 29(11): R447–R450.
Bird R. B., McGuire C., Bird D. W., Price M. H., Zeanah D., Nimmo D. G. 2020. Fire Mosaics and Habitat Choice in Nomadic Foragers. Proceedings of the National Academy of Sciences 117: 12904–12914.
Boehm C. 2012. Moral Origins: The Evolution of Virtue, Altruism, and Shame. New York: Soft Skull Press.
Bradley B. J. 1999. Levels of Selection, Altruism, and Primate Behavior. The Quarterly Review of Biology 74(2): 171–194.
Burkart J. M., Brügger R. K., van Schaik C. P. 2018. Evolutionary Origins of Morality: Insights from Non-human Primates. Frontiers in Sociology 3: 17. URL: https:// doi.org/10.3389/fsoc.2018.00017.
Butovskaya M. L. 2020. Primates as Living Links to Our Past: Variations in Hierarchy Steepness but not Real Egalitarianism. Первобытная археология. Журнал междисциплинарных исследований 1: 13–26.
Butovskaya M. L., Burkova V. N., Karelin D. V. 2020. Does Friendship Matter? Sharing, Fairness and Parochial Altruism in African Children and Adolescents. Social Evolution & History 19(1): 89–112.
Butovskaya M. L., Karelin D. V., Dronova D. A., Filatova V. O., Butovskaya P. R., Tishkov V. A. 2020. Strategies for Sharing Limited Resources among Children and Adolescents in Three Traditional Societies of East Africa: Sociocultural and Genetic Factors. Doklady Biological Sciences 494(1): 219–224.
Butovskaya M., Marczak M., Misiak M., Karelin D., Białek M., Sorokowski P. 2020. Approach to Resource Management and Physical Strength Predict Differences in Helping: Evidence from Two Small-scale Societies. Frontiers in Psychology 11: 373. URL: https://doi.org/10.3389/fpsyg.2020.00373.
Choi J-K. Bowles S. 2007. The Coevolution of Parochial Altruism and War. Science 318: 636–640.
Cronk L, Aktipis A. 2021. Design Principles for Risk-pooling Systems. Nature Human Behavior 5(7): 825–833.
Czárán T., Aanen D. K. 2016. The Early Evolution of Cooperation in Humans. On Cheating, Group Identity and Group Size. Behaviour 153(9–11): 1247–1266.
Fehr E., Bernhard H., Rockenbach, B. 2008. Egalitarianism in Young Children. Nature 454(7208): 1079–1083.
Frayer D. W. 2019. Neanderthals and the Black Swan. PaleoAnthropology 2019: 350–361.
Gintis H., van Schaik C., Boehm, C. 2019. Zoon Politikon: The Evolutionary Origins of Human Socio-political Systems. Behavioural Processes 161: 17–30.
Girard-Buttoz C., Surbeck M., Samuni L., Boesch C., Fruth,B., Crockford C. et al. 2020. Variable Use of Polyadic Grooming and its Effect on Access to Social Partners in Wild Chimpanzees and Bonobos. Animal Behaviour 168: 211–224.
Glowacki L. 2020. The Emergence of Locally Adaptive Institutions: Insights from Traditional Social Structures of East African Pastoralists. Biosystems 198: 104257. URL: https://doi.org/10.1016/j.biosystems.2020.104257.
Glowacki L., Lew-Levy S. 2022. How Small-scale Societies Achieve Large-scale Cooperation. Current Opinion in Psychology 44: 44–48.
Glowacki L., Wrangham R. 2015. Warfare and Reproductive Success in a Tribal Population. Proc Natl Acad Sci Unit States 112: 348–353.
Güroğlu B., Will,G. J., Crone E. A. 2014. Neural Correlates of Advantageous and Disadvantageous Inequity in Sharing Decisions. PloS One 9(9): e107996.
Handley C., Mathew S. 2020. Human Large-scale Cooperation as a Product of Competition between Cultural Groups. Nature Communications 11(1): 1–9.
Henrich J. 2016. The Secret of Our Success: How Culture Is Driving Human Evolution, Domesticating Our Species, and Making Us Smarter. Princeton: Princeton University Press.
Henrich J., Gil-White F. J. 2001. The Evolution of Prestige: Freely Conferred Deference as a Mechanism for Enhancing the Benefits of Cultural Transmission. Evolution and Human Behavior 22(3): 165–196.
Henrich J., Muthukrishna M. 2021. The Origins and Psychology of Human Cooperation. Annual Review of Psychology 72: 207–240.
Hrdy S. 2009. Mothers and Others. The Evolutionary Origins of Mutual Understanding. Cambridge, MA: The Belknap Press of Harvard University Press.
Hrdy S. 2016. Development Plus Social Selection in the Emergence
of “Emotionally Modern” Humans. Childhood: Origins, Evolution, and Implications
/ Ed. by
C. L. Meehan, A. N. Crittenden, pp. 11–44. Albuquerque:
School of Advanced Research Press.
Ishizuka S., Takemoto H., Sakamaki T., Tokuyama N., Toda K., Hashimoto C., Furuichi T. 2020. Comparisons of Between-group Differentiation in Male Kinship between Bonobos and Chimpanzees. Scientific Reports 10(1): 1–8.
Laland K. N., Kumm J., Feldman
M. W. 1995. Gene-culture Coevolutionary Theory:
A Test Case. Current Anthropology 36(1):
131–156.
Lewis L. S., Kano F., Stevens J. M., DuBois J. G., Call J., Krupenye C. 2021. Bonobos and Chimpanzees Preferentially Attend to Familiar Members of the Dominant Sex. Animal Behaviour 177: 193–206.
Mathew S., Boyd R. 2011. Punishment Sustains Large-scale Cooperation in Prestate Warfare. Proceedings of the National Academy of Sciences 108: 11375–11380.
Newson L., Richerson P. 2018. Dual Inheritance Theory. The International Encyclopedia of Anthropology / Ed. by K. D. Keith, pp. 1–5. London: Wiley-Blackwell.
Nolte S., Call J. 2021. Targeted Helping and Cooperation in Zoo-Living Chimpanzees and Bonobos. Royal Society Open Science 8: 201688.
Nowak M. A. 2006. Five Rules for the Evolution of Cooperation. Science 314: 1560–1563.
Pandit S. A., Pradhan G. R., van Schaik C. P. 2020. Why Class Formation Occurs in Humans but not among Other Primates. Human Nature 31(2): 155–173.
Richerson P., Baldini R., Bell A. V., Demps K., Frost K., Hillis V., Mathew S., Newton E. K., Naar N., Newson L., Ross C., Smaldino P. E., Waring T. M., Zefferman M. 2016. Cultural Group Selection Plays an Essential Role in Explaining Human Cooperation: a Sketch of the Evidence. Behavioral and Brain Sciences 39: e30.
Richerson P.J., Boyd R. 2005. Not by Genes Alone: How Culture Transformed Human Evolution. Chicago: University of Chicago Press.
Rodseth L. 2012. From Bachelor Threat to Fraternal Security: Male Associations and Modular Organization in Human Societies. International Journal of Primatology 33(5): 1194–1214.
Samuni L., Crockford C., Wittig R. M. 2021. Group-level Cooperation in Chimpanzees is Shaped by Strong Social Ties. Nature Communications 12(1): 1–10.
Sánchez‐Villagra M. R., Schaik C. P. van 2019. Evaluating the Self‐domestication Hypothesis of Human Evolution. Evolutionary Anthropology: Issues, News, and Reviews 28(3): 133–143.
Santos F. R., Pinotti T., Fujita R. 2021. Population Variation of the Human Genome. Human Genome Structure, Function and Clinical Considerations / Ed. by L. Haddad, pp. 329–350. Cham: Springer.
Sikora M., Seguin-Orlando A., Sousa V. C., Albrechtsen A., Korneliussen T., et al. 2017. Ancient Genomes Show Social and Reproductive Behavior of Early Upper Paleolithic Foragers. Science 358(6363): 659–662.
Sillen-Tullberg B., Bryant E. H. 1983. The Evolution of Aposematic Coloration in Distasteful Prey: an Individual Selection Model. Evolution 37(5): 993–1000.
Singh M., Glowacki L. 2021. Human Social Organization during the Late Pleistocene: Beyond the Nomadic-egalitarian model. EloEvoRxiv Preprints. URL: 10.32942/osf.io/vusye.
Smith K. M., Larroucau T., Mabulla I. A., Apicella C. 2018. Hunter-gatherers Maintain Assortativity in Cooperation Despite High Levels of Residential Change and Mixing. Current Biology 28: 3152–3157.
Tomasello M. 2020a. The Adaptive Origins of Uniquely Human Sociality. Philosophical Transactions of the Royal Society B 375: 20190493.
Tomasello M. 2020b. The Role of Roles in Uniquely Human Cognition and Sociality. Journal for the Theory of Social Behavior 50: 2–19.
Waring T. M., Wood Z. T. 2021. Long-term Gene- Culture Coevolution and the Human Evolutionary Transition. Proceedings of the Royal Society B 288: 20210538.
Whitehead H., Laland K. N., Rendell L., Thorogood R., Whiten A. 2019. The Reach of Gene-Culture Coevolution in Animals. Nature Communications 10(1): 1–10.
Wiessner P. 2002. Hunting, Healing, and Hxaro Exchange: A Long-term Perspective on! Kung (Ju/’hoansi) Large-game Hunting. Evolution and Human Behavior 23(6): 407–436.
Wiessner P. 2019. Collective Action for War and for Peace: A Case Study among the Enga of Papua New Guinea. Current Anthropology 60(2): 224–244.
Wilson E. O. 1978. What is Sociobiology? Society 15(6): 10–14.
Wilson M. L., Boesch C., Fruth B., et al. 2014. Lethal Aggression in Pan is Better Explained by Adaptive Strategies than Human Impacts. Nature 513: 414–417.
Wrangham R. 2017. Control of Fire in the Paleolithic: Evaluating the Cooking Hypothesis. Current Anthropology 58(S16): S303–S313.
Zwir I., Del-Val C., Hintsanen M. et al. 2021. Evolution of Genetic Networks for Human Creativity. Molecular Psychiatry 27: 354–376. URL: https://doi.org/10.1038/s41380-021-01097-y.
* Данная статья подготовлена в рамках гранта РНФ № 18-18-00075.
Для цитирования: Бутовская М. Л. 2022. Эволюция человека: между конкуренцией и сотрудничеством. Эволюция: Эволюционная мозаика / Отв. ред. Л. Е. Гринин, А. В. Коротаев. Волгоград: Учитель. С. 102–119.
For citation: Butovskaya M. L. 2022. Human
Evolution: Between Competition and Cooperation. Evolution: Evolutionary Mosaic / Ed. by L. E. Grinin, A. V.
Korotayev. Volgograd: Uchitel.
Pp. 102–119 (in Russian).




